Introduction
Lead occurs naturally in soil and
water. Plants absorb soil lead through their roots and thus, all plants contain
small amount of this metal (1). The relationship between plants and soil lead
varies with factors ranging from chemical forms of the element in soil, soil
properties, climate, plant species, e.t.c. (2). Lead, a toxic heavy metal and
pollutant of the environment that originates from various sources like mining,
pottery, casting and soldering, metallurgy, aerosols and dust from smelters,
ammunition and automobile-exhaust, gasoline, e.t.c (3). However, increased lead
levels in the soil environment inhibits seed germination, stunts seedling
growth and threatens plant metabolic reactions for proper growth and
development resulting in low yields (4). High levels of heavy metals in the
soil normally result in oxidative damage to plants either directly or
indirectly by triggering an increased level of reactive oxygen species
generation that generally cause damage to the biological molecules such as proteins,
membrane lipids, chloroplast pigments, enzymes, nucleic acids, e.t.c.(5). These
free radicals include superoxide radical (O2*-
) hydroxyl radical (OH*- ) and hydrogen peroxide (H2O2)
that are produced as by products during membrane linked electron transport
reaction and by associated metabolic pathways (6). Oxidative stress occurs
where there is imbalance between the production
of reactive oxygen species and the biological systems ability to readily
detoxify the reactive intermediates produced or failure to easily repair the
damage (7). Also several studies have reported that organosulphur and
polyphenolic compounds in plants protect against oxidative stress (8). To
salvage the rice plants from lead-induced oxidative stress, this study was
designed to assess the uptake and distribution of lead in root and shoot of
rice plants, determine the inhibitory potential of garlic as well as possible
alteration in the activity of some defensive enzymes of rice plants.
Materials and
Methods
Collection
of Soil Sample
Soil samples were collected from
five states in South-West region of Nigeria. Two locations (Toga and Owode) in
Ogun state were considered for the research due to their appreciable lead
levels after a comprehensive soil test.
Preparation
of Garlic Extract
500g of the powdered sample
(garlic cloves) was extracted via maceration for 48hrs using the method of
Aguawa and Mittal (9).
Experimental
Design
In both locations (Toga and Owode
areas), rice seeds were surface sterilized with 0.1% sodium hypochlorite
solution for 10mins and then rinsed with distilled water. After 24hrs
imbibitions of seeds in water, the seedlings were raised in clay (Owode area)
and sand (Toga area) cultures respectively in lead free plastic pots. The pots
received respective treatment solutions and were carefully maintained under
control for seedling growth in a biological oxygen demand (B.O.D) and optimum
relative humidity with 12hrs photoperiod. In each location, the pots were
grouped into four groups A, B, C, and D. A contained seedlings treated with
distilled water only and served as positive control. B seedlings were treated
with 500mg/kg lead acetate only, and seedling in C received 500mg/kg lead
acetate with 500mg/kg garlic extract simultaneously while seedling in D were
treated with 500mg/kg garlic extract only.
Determination
of Lead In Rice Seedlings
Fresh root and shoot samples were
surface sterilized with 1M HCl and then with 1mM Na2EDTA for the
surface bound lead and then dried in oven at 700C for 5-days. Dried
samples were ground to a fine powder in mortar and pestle and digested with
concentrated H2SO4. Digested samples were dissolved in
de-ionized distilled water and lead content was estimated using atomic
absorption spectrophotometer (AAS).
Oxidative
Stress Assay
The level of lipid peroxidation
products was determined using the method of Heath and Packer (10). Fresh root
and shoot samples were ground in 0.25% thiobarbituric acid (TBA) in 10% TCA
using mortar and pestle. The mixture was heated at 950C for 30min
then cooled in an ice bath and centrifuged at 10 000 x g for 10min. the
absorbance of the supernatant was read at 532nm while total of 0.25% TBA in 10%
TCA served as blank. The concentration of lipid peroxides together with the
oxidative-modified proteins of plants were quantified and expressed as total
TBARS as nmol g-1 fresh weight using an extinction coefficient of
155mM-1 cm-1.
Evaluation
of Garlic Inhibitory Potential
The production and inhibition of
lipid peroxides from rice roots and shoots was determined using method
described by (11). The roots and shoots were ground in cold saline (1/10 w/v)
with 10 up-and-down strokes in mortar and pestle. The homogenate was
centrifuged for 10min at 10 000xg to obtain the supernatant and also incubated
with lead acetate and garlic extract at varied concentrations together with
de-ionized water at total volume 300µl at 370C for 1hr. The color
reaction was monitored by adding 200, 250 and 500µl each of 8.1% sodium
dodecylsulphate (SDS), acetic acid at pH 3.4 and 0.6% TBA respectively. The
solution was incubated at 970C for 1hr and absorbance was read at
532nm.
Superoxide
dismutase Assay
The activity of superoxide
dismutase (SOD) was determined according to method described by (12). About 200
mg fresh tissue (root/shoot) were homogenized in 5ml of 100 mM K-phosphate
buffer at pH 7.8 containing 0.1 mM EDTA. 0.1% (v/v) Triton X-100 and 2% (w/v)
polyvinyl pyrrolidone (PVP). The extract was filtered and centrifuged at 22 000
x g for 10mins at 4 0C. The supernatant was dialyzed in cellophane
membrane tubings against the cold extraction buffer for 4hrs with 3-4 changes
of the buffer and later used for the assay. The assay mixture in a total volume
of 3ml contained 50 mM sodium carbonate-bicarbonate buffer (pH 9.8), 0.1 mM
EDTA, 0.6mM epinephrine and enzyme while epinephrine was added last. The
adrenochrome formation after 5mins was recorded at 475nm in a UV-Vis
spectrophotometer.
Catalase
Assay
The activity of catalase was
examined according to (13). 200 mg Fresh tissue (roots/shoot) were homogenized
in 5ml of 50 mM EDTA. 2% (w/v) PVP and 0.5% (v/v) Triton X -100. The homogenate
was centrifuged at 22 000 x g for 10mins at 40C and after which the
supernatant was used for the enzyme assay. The assay mixture in total volume of
1.5ml contained 1000µl of 100µl enzyme at 240 nm.
Glutathione
reductase Assay
Glutathione reductase was assayed
according to (14). 200mg Fresh tissue (root/shoot) were homogenized using
chilled mortar and pestle in 5ml of 50 mM Tris-HCl buffer (pH 7.6). The
homogenate was centrifuged at 22 000 x g for 30mins at 4 0C and the
supernatant was used for the enzyme. The reaction mixture in a total volume of
1ml contained 50 mM Tris-HCl buffer (pH 7.6), 0.15 mM NADPH, 1 mM GSSG, 3 mM
MgCl2 and 200µl enzyme extract. The activity of the enzyme was
monitored with absorbance at 340nm.
Results
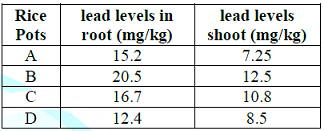
Table 1: Uptake of lead by growing rice seedlings (Sand) in Toga area for 40-days.
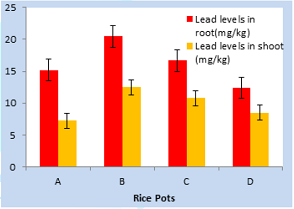
Figure 1: Uptake of lead by growing rice seedlings (Sand) in Toga area for 40-days.
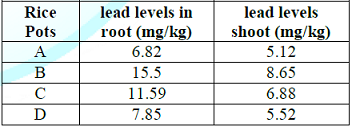
Table 2: Uptake of lead by growing rice seedlings (clay) in Owode area for 40-days.
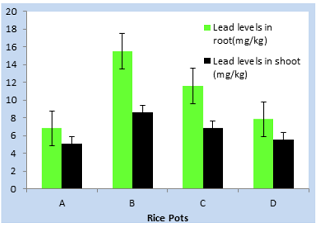
Figure 2: Uptake of lead by growing rice seedlings (clay) in Owode area for 40-days.
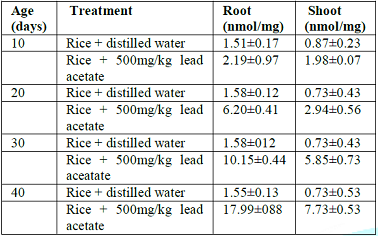
Table 3: Levels of total lipid peroxides in root and shoot of rice seedlings in Toga area.
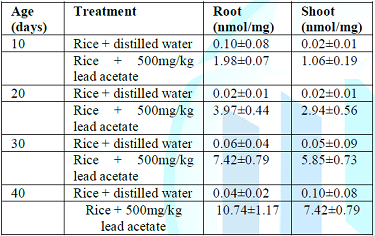
Table 4: Levels of total lipid peroxides in root and shoot of rice seedlings in Owode area.
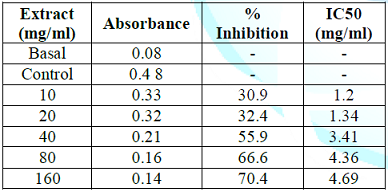
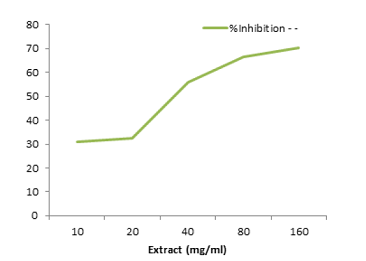
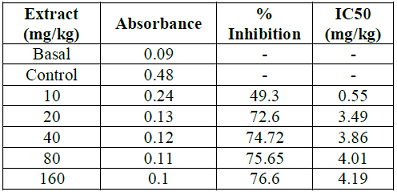
Table 5: Inhibitory potential of garlic aqueous extract on lead acetate induced oxidative stress in root of rice plant.
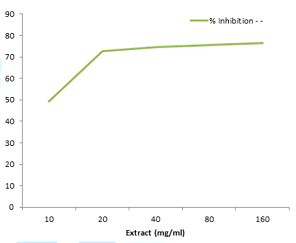
Figure 3: Inhibitory potential of garlic aqueous extract on lead acetate induced oxidative stress in root of rice plant.
Table 6: Inhibitory potential of garlic aqueous extract on lead acetate induced oxidative stress in shoot of rice plant.
Figure 4: Inhibitory potential of garlic aqueous extract on lead acetate induced oxidative stress in shoot of rice plant.
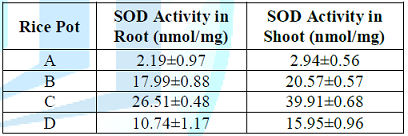
Table 7: Effect of lead uptake on superoxide dismutase activity in rice seedlings.
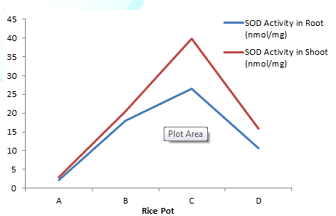
Figure 5: Effect of lead uptake on superoxide dismutase activity in rice seedlings.
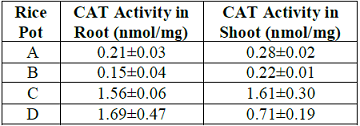
Table 8: Effect of lead uptake on catalase activity in rice seedlings.
Figure 6: Effect of lead uptake on catalase activity in rice seedlings.
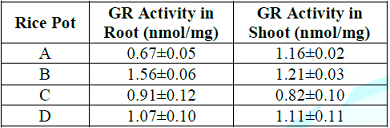
Table 9: Effect of lead uptake on glutathione reductase activity in rice seedlings.
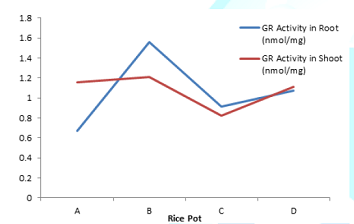
Figure 7: Effect of lead uptake on glutathione reductase activity in rice seedlings.
Discussion
Lead uptake highest level
(20.50mg/kg) was observed in root treated with 500mg/kg lead acetate in pot B.
lowest lead level was obtained in pot D in root treated with garlic extract
only in Toga area for 40-days. However, highest lead level obtained in shoot
(12.50mg/kg) in pot B treated with lead acetate only was much more reduced
compared to root, while lowest lead level (8.50mg/kg) was obtained in D. This
was reasonably possible as plant roots normally absorb more lead than shoot. In
addition at Owode area, (15.50mg/kg) and (8.65) were the highest and lowest
lead levels obtained in rice root and shoot respectively. These values were
comparatively lower to the values of lead concentrations obtained for both root
and shoot of rice plant in Toga area. This could be attributed to enhanced lead
absorption aided by wet or flooded soils in Toga area. However, due to
antioxidant property of garlic, lead absorption was restricted in pot D where
remarkable lowest lead level was obtained. Besides, the production of lipid
peroxides in both root and shoot of rice seedling showed increased lead
concentration relative to seedling age or period of growth. Seedlings grown
under 10-days had (1.5nmol/mg and 0.87nmol/mg) lowest concentration of lipid
peroxides in rice root and shoot respectively. Those grown under 40-days had
(17.99nmol/mg and 7.73nmol/mg) highest concentration of lipid peroxides in rice
root and shoot respectively in Toga swampy area. However, same trend was
observed in Owode area where highest lipid peroxidation levels (10.74nmol/mg
and 7.42nmol/mg) in root and shoot respectively were obtained under 40-days
while lowest lipid peroxidation levels (0.10nmol/mg and 0.02nmol/mg) in root
and shoot were respectively obtained under 10-days.
In view of the above, peroxide
levels in Toga area was higher than that obtained in Owode area and maximum
concentration (17.99nmol/mg) was obtained in rice root. This is because root
absorbs lead faster in wet or flooded area and tends to retain absorbed lead
which it does not easily transport for prompt uptake by the plant shoot. This
results to slow mobility of lead in rice plant where large accumulations are
observed in root. The high lipid peroxide levels in root due toaccumulation
could as well be attributed to the ability of Pb2+ which catalyzes
one electron (e-) transfer reaction that generate reactive oxygen species (ROS)
such as hydroxyl radicals and hydrogen peroxides via Fenton reaction and thus,
generate more lipid peroxides (15). However, the protective effect demonstrated
by garlic in this study could be due to presence of its inherent phenolic and
organosulphur components that constitute and enhance its antioxidant activity.
The antioxidant activity has been
reported to be concomitant with the development of reducing power (16) and this
is due to garlic extract hydrogen donating ability (17). In this study, garlic antioxidant
activity was demonstrated against the reactive oxygen species generated and
thus, inhibits lipid peroxidation due to its scavenging potential. Besides,
plants generally possess inherent antioxidant defense system used naturally to
combat the oxidative damage. In view of this, table 7 shows effect of lead
uptake on Superoxide-dimutase (SOD) activity in rice seedling where a
significant (P<0.05) increase in SOD activity (26.51 and 17.99) was observed
in roots of rice seedlings of pots C and B treated with lead acetate.
Meanwhile similar effect was
observed in shoots of rice seedlings of pots C and D treated with lead acetate
where significant (P<0.05) increase (39.91 and 20.57) in SOD activity
respectively. However, SOD activity was observed higher in shoot than root
because root absorbs more lead concentrations than shoot. Hence, the excessive
lead has the propensity of reducing the defensive potential of SOD in plant
root. SOD activity has been reported to increase under water stress (18), heavy
metal toxicity (19). This increase in response to stress could be due to de
novo synthesis of the enzyme (20). The catalase (CAT) activity observed for
lead treated rice seedlings in pot B was significantly (P<0.05) lower than
control for both root (0.15) and shoot (0.22) respectively. However, in pots C
and D treated with the sample extract, CAT activity was appreciably higher in
the plant tissues when compared to control. The decline in CAT activity in
pot B could be attributed to lead toxicity which could possibly delay the
removal of hydrogen peroxide and peroxides mediated by catalase which in turn enhances
free radical mediated lipid peroxidation in plant tissues (21). Besides, a
decline in catalase activity has been attributed to the inactivation of enzyme
protein due to deleterious activity of reactive oxygen species which either
decrease the enzyme synthesis or cause alteration in assembly of enzyme
subunits (22). On the contrary, glutathione reductase (GR) activity was higher
in both root (1.56) and shoot (1.21) of rice seedlings treated with lead
acetate in pot B compared with control. Similarly, GR activity was
significantly (P<0.05) higher in pot D treated with extract sample than
tissues of rice seedlings in pot C. This could be attributed to antioxidant
potential of the extract (garlic) which also compliments the GR antioxidant
activity (23). In addition, the increased GR activity suggests possible
involvement of GR in regenerating GSH from GSSG under lead toxicity in order to
increase GSH/GSSG ratio and thus, increasing total glutathione pool (24). The
study above clearly suggests that lead toxicity induces oxidative stress in
rice plants which could be modulated by garlic antioxidant effect, while
antioxidant enzymes play a pivotal role in combating oxidative stress in the
plants
Acknowledgement
The authors
appreciated the financial aid and facilities provided by tertiary education
trust fund (TETFUND) via Federal Government of Nigeria.
References
1.
Erik MI, Peak JD, Brady PV and Pesek
JD. Kinetics of lead absorption and deposition in soil (1999) Soil Sci 64:
28-39.
2.
Godbold DI and Kettner C. Lead
influence on root growth and mineral nutrition of Picea abies seedlinds (1991)
J Plant Physiol 139: 95-99. http://dx.doi.org/10.1016/S0176-1617(11)80172-0
3.
Mayer DT and Brown K. Roles of
peroxidase in development of water impermeable seeds (2008) J Phys. Biochem
193: 265-275. https://doi.org/10.1007/BF00405186
4.
Kasfori R and Petrovic M. Effect
of excess lead, cadmium copper and zinc on water relation in sunflower (2008) J
Plant Nutri 15: 2427-2439. https://doi.org/10.1080/01904169209364485
5.
Malecka A, Jarmuseklewiez W and Tomaszewska
B. Antioxidative defense against lead induced oxidative stress in some cellular
components of pea root cells (2001) Acta Biochem 48: 687-698.
6.
Becana M, Dalton DA, Moran JF and
Matamoros MA. Reactive oxygen species and antioxidants in legume nodules (2000)
Plant Physiol 109: 372-381. https://doi.org/10.1034/j.1399-3054.2000.100402.x
7.
Halliwell B. Free radicals and
antioxidant defense system (1994) J Lab Clin Med 119: 598-620.
8.
Nwanjo HU. Free radical
scavenging potential of the aqueous extract of Viscum album (mistletoe) leaves
(2007) Int J Nutrition wellness 12: 121-127.
9.
Aguawa CN and Mittal GC. Study of
the antioxidative potentials of Pyrynacanthia staudtii using various models of
experiments (1981) Envr J Pharmacol 40: 215-220.
10. Heath
RI and Packer L. Photoperoxidation in isolated chloroplast; kinetics and
stochiometry of fatty acid peroxidation (1968) Arch Biochem Biophys 125:
189-198. https://doi.org/10.1016/0003-9861(68)90654-1
11. Ohkawa
H, Ohishi N and Yagi K. Assay for lipid peroxides via thiobarbituric acid
reactions (1979) Analytical Biochem 95: 351-358. Misra HP and Fridovich I. The
role of superoxide anion in auto-oxidation of epinephrine and simple assay for
superoxide dismutase (1972) J Biol Chem 247: 3170-3175.
12. Beers
RF and Sizer IW. Colorimetric method for estimation of catalase (1982) J Biol
Chem 195: 133-139.
13. Schaedle
M and Bassham JA. Chloroplast glutathione-reductase (1997) Plant Physiol 59:
1011-1012. https://doi.org/10.1104/pp.59.5.1011
14. Zago
MP, Verteraesten SV and Oteiza PI. Zinc in the prevention of Fe2+ initiated
lipid and protein oxidation (2000) Biological Res 33: 143-150.
15. Tanaka
T, Kato T, Nishioka Y and Kimura J. Lead burden and oxidative stress effect
(1985) Biol Chem 263: 11646-11651.
16. Baisak
R, Rana DA, Kar M and Acharya PBB. Alteration in the activities of active
oxygen scavenging enzymes of wheat leaves subjected to water stress (1994)
Plant Cell Physiol 35: 489-495. http://dx.doi.org/10.1093/oxfordjournals.pcp.a078620
17. Zhang
J and Kirkham MB. Drought stress induced changes in the activities of
superoxide dismutase, catalase and peroxidase in wheat species (1994) Plant
Cell Physiol 35: 785-791. https://doi.org/10.1093/oxfordjournals.pcp.a078658
18. Shah
K, Kumar RG, Verma S and Dubey RS. Effect of cadmium on lipid peroxidation,
superoxide anion generation and activities of antioxidant enzymes in growing
rice seedlings (2001) Plant Sci 161: 1135-1144. https://doi.org/10.1016/S0168-9452(01)00517-9
19. Lozano
R, Azoon R and Palm JM. Superoxide dismutase and drought stress in lactna
sativa (1996) New Phytol 136: 329-333.
20. Lin
CC and Kao CH. Effect of sodium chloride stress on hydrogen peroxide metabolism
in rice leaves (2000) Plant Growth Regul 30: 151-155. https://doi.org/10.1023/A:1006345126589
21. Mac
Rae EA and Ferguson IB. Changes in catalase activity and hydrogen peroxide
concentration in plants in response to low temperature (1985) Plant Physiol 65:
51-56. https://doi.org/10.1111/j.1399-3054.1985.tb02358.x
22.
Scandalios JG. Response of plant
antioxidant defense genes to environmental stress (1990) Advance Genet 28:
11-41. https://doi.org/10.1016/S0065-2660(08)60522-2
23. Noctor G and Foyer CH. Ascorbate and
glutathione; keeping active oxygen under control (1998) Annual Reviews Plant
Mol Biol 499: 249-279. https://doi.org/10.1146/annurev.arplant.49.1.249
*Corresponding author:
Tugbobo
Oladimeji S, Associate Professor of Biochemistry, Toxicology and Plant
Biochemistry Unit, Department of Science Technology, Federal Polytechnic,
Ado-Ekiti, Nigeria, Tel: +2348035303701, Email: tugbobooladimeji1@gmail.com
Citation:
Tugbobo OS, Idowu KS and Oluwaseyi AI. Antioxidative
potential of garlic on lead-induced oxidative stress and effect on enzyme
activity in rice plants (2018) Edelweiss Appli Sci Tech 2: 79-83




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}